Mike Stone: Nedostatek kontroly ze strany virologie

Přeložil: Vladimír Bartoš

V průběhu let se vedlo mnoho diskusí o tom, zda virologie dodržuje vědeckou metodu, či nikoliv, přičemž většina těchto diskusí se zaměřovala na nedostatek řádných kontrol. Spor se soustředil na to, zda kontroly, které se někdy používají, ale jsou zřídka popsány, známé jako falešné infekce (mock infections), jsou vůbec správnou kontrolou. Pro ty, kteří nejsou obeznámeni, vědecké kontroly jsou kontrolním a vyvažovacím systémem, který se používá během experimentů, když se výzkumníci snaží určit příčinu nějakého účinku. Kontroly jsou navrženy tak, aby zajistily, že předpokládaná příčina, známá jako nezávislá proměnná (IV), je jedinou věcí, která by mohla způsobit pozorovaný účinek, známý jako závislá proměnná (DV):
“Studie s kontrolou (kontrolami) je navržena tak, aby zajistila, že účinky jsou způsobeny nezávislými proměnnými v experimentu. Použití kontrol umožňuje studovat vždy jednu proměnnou nebo faktor. Je však důležité, aby jak kontrolní, tak druhá (experimentální) skupina (skupiny) byly vystaveny stejným podmínkám kromě jedné zkoumané proměnné. Takový postup pomůže vyvodit přesnější a spolehlivější závěry.”
https://www.biologyonline.com/dictionary/control
Jedním z hlavních důvodů, proč jsem se ve skutečnosti tolik nevěnoval nedostatku platných kontrol ve virologii, je to, že existují jiné, mnohem naléhavější problémy, které je třeba řešit nejdříve, než se vůbec dostaneme do experimentální fáze, abychom mohli diskutovat o řádných kontrolách. Podívejte se na chvíli na jednotlivé kroky vědecké metody:
- Pozorujte jev
- Vytvoření alternativní hypotézy
- Nezávislá proměnná IV (předpokládaná příčina)
- Závislá proměnná DV (pozorovaný účinek)
- Kontrolní proměnné
- Nulová hypotéza
- Test/experiment
- Analýza pozorování/údajů
- Potvrzení/nepotvrzení hypotézy
Určení IV, DV a kontrolních proměnných přichází po pozorování přírodního jevu a stanovuje se při tvorbě hypotézy. Virologové nemohou dokončit ani první krok vědecké metody, protože nejsou schopni pozorovat žádné přírodní jevy. Nejsou schopni vidět předpokládané částice “viru” v přírodě. Nemohou být svědky toho, jak se tyto částice dostanou do člověka a způsobí nemoc, ani nemohou vidět, jak se tyto částice přenášejí z člověka na člověka a způsobují nemoc. Bez pozorování přírodního jevu není možné stanovit platnou hypotézu, aby bylo možné navrhnout experiment, který by ověřil příčinu pozorovaného jevu. To znamená, že krok číslo 2 vědecké metody je rovněž mimo hru. S neschopností dosáhnout prvních dvou kroků vědecké metody se vytrácí i schopnost navrhnout a provést vědecký experiment. Spor o to, zda jsou kontroly používané při pseudovědeckých pokusech s buněčnými kulturami platné, či nikoliv, je tedy příliš předčasný. Celá premisa samotných pokusů s buněčnými kulturami je neplatná, jak ukázal člověk, který tuto metodu vytvořil.
Vytvoření buněčné kultury
“Až do roku 1952 se virologové domnívali, že virus je toxická bílkovina nebo enzym, který přímo otravuje organismus, a že se nějakým způsobem v těle sám množí a šíří se v těle i mezi lidmi a mezi zvířaty. Medicína a věda se této myšlenky vzdaly v roce 1951, protože podezřelý virus nebyl nikdy spatřen v elektronovém mikroskopu a především nebyly nikdy provedeny žádné kontrolní pokusy. Bylo uznáno, že i ze zdravých zvířat, orgánů a tkání se během rozkladu uvolňují stejné produkty tkáňového rozkladu, které byly dříve mylně interpretovány jako “viry”. Virologie vyvrátila sama sebe.”
Dr. Stefan Lanka https://viroliegyhome.files.wordpress.com/2022/08/wissenschafftplus-the-virus-misconception-part-1.pdf
(ZDE CZ verze textu)
Na počátku 50. let 20. století byla virologie na smrtelné posteli poté, co po desetiletí neúspěšně zkoušela purifikovat a izolovat domnělé “virové” částice, aby přímo prokázala existenci a patogenitu těchto neviditelných entit. Za všechna léta pokusů neměli virologové k dispozici nic než nepřímé důkazy rozkladu z pokusů s lidskými a zvířecími tkáňovými kulturami, o nichž se tvrdilo, že jsou způsobeny dotyčným “virem”.

Pasteurův ústav, Kasauli, Indie: jedna fáze přípravy vakcíny proti vzteklině: králičí mozek na čtverci mušelínu. Fotografie, asi 1910.
Výsledky pokusů však nebyly specifické pro přítomnost nějakého předpokládaného “viru”, protože stejný stav rozpadu byl pozorován i u tkání zdravých hostitelů. Jak upozornil Dr. Stefan Lanka ve výše uvedeném citátu, virologie vyvrátila své vlastní pseudovědecké pokusy s tkáňovými kulturami. Navíc zjištění účinků spojených s určitým “virem” od jedné skupiny výzkumníků nemohla být reprodukována ani zopakována jinými skupinami výzkumníků. Ve skutečnosti byly výsledky často v rozporu s tím, co bylo považováno za zavedené důkazy. Výzkumníci se dokonce nemohli shodnout ani na tom, co přesně je “virus”, ani na tom, zda jsou nabízené experimentální důkazy skutečně platné, či nikoli. Skvělé shrnutí tohoto rozkolu podal v roce 1999 Karlheinz Lüdtke ve své eseji:
On the history of early virus research (K historii raného výzkumu virů).
“S “filtrovatelným” virem bylo objeveno něco, co podle tradičních koncepcí, které se koneckonců většinou osvědčily při výzkumu infekčních chorob, nebylo možné popsat způsobem, který by mohli sdílet všichni badatelé. Vznikly velmi rozdílné interpretace podstaty tohoto jevu, které byly stavěny proti sobě. Žádná strana nemohla předložit experimentální důkazy pro tu či onu koncepci, kterou by měli všichni badatelé přijmout. Jinými slovy, rozhodnutí o tom, zda to či ono vysvětlení nejpřesněji vyjadřuje “pravou” povahu viru, nebylo možné empiricky “objektivizovat”. Každá verze výkladu jevu zůstávala otevřená útokům, fakta předkládaná odborné veřejnosti mohla být často oponenty reinterpretována ve fikce, které jako zdroje chyb uváděly do hry závislost zjištění na podmínkách pozorování, lokální situaci experimentů, výzkumnou povahu atribucí vlastností atd. Například zjištění, která často uváděli někteří tehdejší výzkumníci viru, nebyla potvrzena jinými výzkumníky jako výsledek jejich vlastních experimentů, nebo pozorování nemohli reprodukovat všichni vědci, kteří s virem pracovali. Často byly hlášeny opačné nálezy nebo byly zkoumané nálezy považovány za artefakty. Stejně jako v případě zdůvodnění bylo možné se odvolávat na důvody různého druhu, aby bylo možné diskutovaná stanoviska odmítnout. K nálezům, které sloužily k empirickému potvrzení domnělé souvislosti, se často brzy přidaly negativní nálezy hlášené jinými badateli. Jakkoli pečlivě a promyšleně byly při experimentech používány vhodné techniky a navzdory tomu, že každá strana mohla nabídnout věrohodné důvody pro obhajobu svých pozic a předložit empirické důkazy – což vysvětluje, proč “různí oponenti ‘konstruovali’ značně odlišné výzkumné objekty, které označovali za ‘virus'” (van Helvoort 1994a: 202) -, v žádném okamžiku nenabídli přesvědčivé důvody, které by druhou stranu přiměly k tomu, aby se obvinění z artefaktů nakonec vzdala.” (van Helvoort 1994a: 202)
Lütke HISTORIE RANÉHO VÝZKUMU VIRŮ _ENG Stáhnout
Zatímco se v oboru očividně nedařilo, virologii se dostalo podpory v podobě nové metody, která měla prokázat existenci těchto neviditelných entit. Jednalo se o metodu buněčných kultur, kterou v roce 1954 zavedl John Franklin Enders při svých pokusech o identifikaci “viru” spalniček. Protože virologové stále nemohli řádně purifikovat ani izolovat předpokládané částice “viru” přímo z tekutin nemocného hostitele, bylo rozhodnuto, že částice musí být místo toho vypěstovány v kultuře, protože se tvrdilo, že v tekutinách není přítomen dostatek částic. “Virus”, který nemohl být nalezen přímo v tekutinách, aby mohl být řádně studován, nějakým způsobem potřeboval hostitelskou buňku, aby se mohl replikovat a aby mohl být nalezen a studován. Metoda buněčných kultur prý poskytovala údajně lepší nepřímé důkazy než ty předchozí a v konečném důsledku oživila skomírající obor. Aby Enders, virolog, který získal Nobelovu cenu (rovněž v roce 1954) za objevení schopnosti “virů” poliomyelitidy růst v různých typech tkáňových kultur, dosáhl tohoto lepšího nepřímého důkazu, nahradil zvířecí a lidské tkáňové kultury zvířecími a lidskými buněčnými kulturami. Jinými slovy, Enders byl ironicky oceněn Nobelovou cenou za důkazy, které shromáždil pomocí starých vyvrácených postupů tkáňových kultur, které byly následně nahrazeny jeho novou metodou buněčných kultur ještě v témže roce.

Když Enders vytvořil svou metodu buněčných kultur, je třeba poznamenat, že stále ještě nebylo dosaženo prvního a druhého kroku vědecké metody, takže se vrhl do experimentování, aniž by pozoroval přírodní jev, určil závislou proměnnou (tj. účinek), ani neizoloval nezávislou proměnnou (tj. “virus”), aby mohl stanovit hypotézu, kterou by testoval. To však nezabránilo jeho snaze vytvářet pseudovědecké experimenty, jejichž cílem bylo přesvědčit svět, že tyto fiktivní entity existují. I když jsem se zde velmi podrobně zabýval neplatnou praxí buněčných kultur, poskytnu stručné vysvětlení toho, v čem Endersova metoda spočívala. Ve své zásadní práci o spalničkách Enders odebral výplachy z krku pacientů s podezřením na spalničky (které získal v kloktaném odtučněném mléce) a přidal tyto vzorky k lidským a opičím ledvinovým buňkám. Do kultury přimíchal hovězí plodovou vodu, extrakt z hovězího embrya, koňské sérum, antibiotika, inhibitor sójového trypsinu a fenolovou červeň jako indikátor buněčného metabolismu. Tuto směs pak inkuboval několik dní a tekutiny pasážoval 4. a 16. den. Enders nakonec pozoroval tzv. cytopatogenní efekt, což je vzorec poškození, který se v kultuře objevuje, když se buňka rozpadá a odumírá. Enders předpokládal, že tento účinek je přímým důsledkem neviditelného “viru” uvnitř výplachů z odtučněného mléka v krku, který proniká do buňky prostřednictvím lýzy membrány buněčné stěny, což má za následek smrt buňky a replikaci “viru”. Jinými slovy, předpokládal, že buněčné zbytky z otrávené buňky nejsou rozbité kusy kdysi neporušené buňky, ale jsou to nově vytvořené “virové” kopie. Navzdory nevědecké povaze této metody se buněčná kultura rychle prosadila jako “zlatý standard” pro “izolaci viru” a virologové ji používají dodnes:
“Buněčná kultura poskytuje optimální prostředí pro detekci a identifikaci četných lidských patogenů, čehož se dosahuje prostřednictvím izolace virů v buněčné kultuře jako “zlatého standardu” pro objevování virů.”
Je třeba si uvědomit, že spolu s tím, že Enders neměl platnou nezávislou proměnnou ve formě purifikovaných a izolovaných “virových” částic, vytvořil svou vlastní závislou proměnnou v cytopatogenním účinku. Tento účinek není přirozeně pozorovaným jevem. Lze jej pozorovat pouze prostřednictvím experimentů a manipulací v laboratoři. Enders již předpokládal, že “virus” existuje a že bude pozorovat účinek, pokud přidá vzorek obsahující předpokládaný “virus” do buněčné kultury. Jakmile byl svědkem tohoto efektu, Enders tvrdil, že se jedná o přímý důsledek přítomnosti “viru”, přestože “virus” v buněčné kultuře neviděl. Tím se Enders dopustil tzv. logického klamu, který se obvykle vyjadřuje takto: “Tvrzení následku” (affirming the consequent logical fallacy):

To je také příkladem kladení otázek a argumentace kruhem, jak krásně ilustruje tento graf Aleca Zecka a Dr. Andrewa Kaufmana:

Z toho je patrné, že Endersova metoda je nejen nevědecká, protože se neřídí vědeckou metodou, ale jeho závěry jsou také plné logických chyb. To však nebyly jediné problémy Endersovy experimentální metody.
Enders vyvrátil sám sebe
Přestože byla buněčná kultura přijata jako “zlatý standard” důkazu pro “izolaci” “viru”, pravidelně se opomíjí zmínit jedna věc: Enders sám si nebyl jistý, zda je jeho metoda vůbec platná. Ve své práci z roku 1954 Enders zpochybnil, zda lze říci, že výsledky pokusů vytvořené v laboratoři (in vitro) odrážejí to, co se děje uvnitř těla (in vivo):
“Patologické změny vyvolané těmito látkami v epiteliálních buňkách v tkáňové kultuře se alespoň povrchně podobají těm, které se vyskytují v některých tkáních během akutní fáze spalniček. Ačkoli není žádný důvod k závěru, že faktory in vivo jsou stejné jako ty, které jsou základem tvorby obrovských buněk a poruch jádra in vitro, výskyt těchto jevů v kultivovaných buňkách odpovídá vlastnostem, které by a priori mohly být spojovány s virem spalniček.“
Enders rovněž připustil, že jeho nepřímé důkazy, které použil pro spojení s neviditelným “virem” spalniček, byly neúplné:
“Ačkoli jsme tedy již získali značné nepřímé důkazy podporující etiologickou roli této skupiny původců spalniček, zbývá provést ještě 2 experimenty, které jsou pro stanovení tohoto vztahu zásadní.”
Nejvíce usvědčujícím odhalením však bylo přiznání, že během svých experimentů s předpokládaným “virem” spalniček pomocí metody buněčných kultur, kterou vynalezl, pozoroval Enders přesně tytéž cytopatogenní účinky, které spojoval s “virem” spalniček, i v normálních kontrolních kulturách bez přítomnosti jakéhokoli “viru”:
“Kultury opičích ledvin lze tedy použít ke studiu těchto agens stejným způsobem jako kultury lidských ledvin. Přitom je však třeba mít na paměti, že cytopatické účinky, které se povrchně podobají účinkům infekce původci spalniček, mohou být případně vyvolány jinými virovými agens přítomnými v tkáni opičích ledvin (viz poslední odstavec v části G) nebo neznámými faktory.”
“Druhé agens bylo získáno z neinokulované kultury buněk opičích ledvin. Cytopatické změny, které vyvolal v neobarvených preparátech, nebylo možné s jistotou odlišit od virů izolovaných ze spalniček. Když však byly buňky z infikovaných kultur fixovány a obarveny, bylo možné jejich účinek snadno odlišit, protože nebyly pozorovány internukleární změny typické pro původce spalniček. Navíc, jak jsme již uvedli, tekutiny z kultur infikovaných původcem nefixovaly komplement v přítomnosti séra rekonvalescentů spalniček. Je zřejmé, že možnost setkání s takovými agens ve studiích se spalničkami je třeba mít neustále na paměti.”
Jinými slovy, John Franklin Enders pomocí neočkovaných kultur zjistil, že cytopatogenní účinek, o němž předpokládal, že je způsoben neviditelným “virem” spalniček, se objevil i tehdy, když v kultuře žádný předpokládaný “virus” nebyl přítomen. Byl to tedy samotný proces kultivace buněk zahrnující hladovění buňky s minimem živin a otravu buňky toxickými chemikáliemi, přísadami a cizorodými látkami, které nakonec vedly k odumření buňky a následnému cytopatogennímu účinku. Proto by se mělo dojít k závěru, že cytopatogenní účinek není dílem žádného fiktivního “viru”. Toto odhalení mělo virologii ukončit právě tehdy.
Místo toho však byly výsledky Endersovy kontroly zameteny pod koberec a ignorovány. Místo toho, aby virologii odpískali konec, jak měl učinit každý, kdo má alespoň špetku intelektuální poctivosti, odpovědní činitelé zdvojnásobili neplatnou metodu buněčných kultur jako standard, který musí dodržovat každý virolog, aby potvrdil přítomnost “viru” ve vzorku. Toto spoléhání se na důkaz buněčnou kulturou postavený na podvodném cytopatogenním základu ještě více zabetonovalo celý obor virologie do šíleného světa pseudovědy, protože vědecká metoda byla nadále opouštěna.
Enders vyvrácen jinými

Zajímavé je, že když se další vědci snažili obnovit Endersovy výsledky, stále se objevovaly chyby v metodice. Ve skutečnosti různé týmy výzkumníků během následujících pěti let zjistily naprosto stejné cytopatogenní výsledky jako Enders, když si sami prováděli neinokulované kontroly. Dr. Cowan udělal vynikající přehled tří z těchto studií v tomto nedávném videu:
Podívejme se na výňatky ze tří prací, o nichž Dr. Cowan hovořil, abychom viděli, co přesně tito výzkumníci zjistili. Zařadil jsem do nich nejdůležitější momenty z každé práce a také části o metodách, které ukazují absurdní kultivační kroky, které každý tým použil, aby se pokusil “izolovat” své “viry”.
V této první studii z roku 1955 od Rustigiana a spol. se uvádí, že výzkumníci objevili neidentifikovaný “agens” v rámci svých neinokulovaných kontrol při pokusu o adaptaci “viru” dengue na válečkové zkumavkové kultury ledvin opic rhesus. Tento “agent” vyvolal přesně stejný cytopatogenní účinek, jaký pozoroval Enders u své neinokulované kultury během svých pokusů se spalničkami. Po přípravě kultur opičích ledvin pro studii poliomyelitidy se v neinokulovaných kulturách objevily další 3 neidentifikované “agens”, které vyvolaly naprosto stejný cytopatogenní účinek. Zdrojem těchto ledvinových kultur byly zdravé opice nevykazující žádné příznaky onemocnění, a proto se nepředpokládalo, že by v kultuře byl přítomen nějaký “virus”. Po zkoumání, zda pozorovaný CPE mohl pocházet z média, dospěli vědci k závěru, že v buňkách zdravých opičích ledvin bylo přítomno něco, co vyvolalo cytopatogenní účinek:
Infekce tkáňových kultur opičích ledvin virovými agens.
“Široký současný zájem o tkáňové kultury pro studium vztahů mezi viry a hostitelskými buňkami a aplikované problémy virových onemocnění zdůraznil potřebu používat buňky a tkáně prosté mikrobiálních agens. Nedávno bylo zdůrazněno nebezpečí vnášení virů do tkáňových kultur s kontaminovanými médii (1). Stejně důležitý, ne-li důležitější, je problém přítomnosti virů v tkáňových kulturách v důsledku nerozpoznané infekce buněk a tkání použitých jako primární explantáty. Detekce takovýchto virových infekcí může být v některých případech komplikována skutečností, že se neprojevují žádné snadno rozpoznatelné změny. Na druhé straně mohou některé viry za stejných podmínek následně výrazně zasahovat do buněčného růstu, což se projevuje inhibicí produkce kyselin nebo cytologickými změnami.
Při našich pokusech v roce 1953 adaptovat havajský virus dengue(2) adaptovaný na myši na válečkových zkumavkách s ledvinami opice rhesus jsme narazili na neidentifikovaný agens, který vyvolal cytopatogenní změny v kulturách opičích ledvin a rakovinných epiteliálních buněk HeLa. Tento agens byl filtrovatelný a nemohl být kultivován v různých neživých médiích. Následně byly z neinokulovaných kultur opičích ledvin připravených pro studie poliomyelitidy do buněčných kultur HeLa přeneseny další 3 látky s identickými cytopatogenními vlastnostmi. Enders a Peebles(3) nedávno informovali o získání agens z neočkované kultury opičích ledvin, které má zřejmě stejné cytopatogenní vlastnosti v opičích ledvinových kulturách jako naše agens. Kromě popisu jeho cytopatického účinku v opičích ledvinových kulturách však neuvedli žádné další údaje týkající se jejich agens. Vzhledem k širokému používání opičích ledvinových kultur při studiích virů se zdá důležité uvést naše pozorování s těmito činiteli. Ta zahrnují jejich cytopatický účinek v ledvinové tkáni opic rhesus, v HeLa buněčných a některých dalších tkáňových kulturách, způsob jejich obnovy a pasáže v tkáňové kultuře a další charakteristiky. Kromě toho jsou uvedeny důkazy, že zdrojem těchto látek v tkáňových kulturách opičích ledvin jsou ledviny zjevně zdravých opic, a nikoliv součásti média.”
Materiály a metody. Technika tkáňových kultur. A. Válečkové zkumavkové kultury. Při přípravě válečkových zkumavkových kultur byly v zásadě dodrženy postupy Robbinse, Wellera a Enderse (4). Kromě ledvin opice rhesus byly připraveny kultury z opičích varlat, lidských embryonálních ledvin, lidské embryonální kůže a svaloviny a ledvin myších mláďat. V prvních studiích se kultivační médium skládalo z Hanks-Simmsova roztoku (70 %) , extraktu hovězího embrya (10 %) a koňského séra (20 %). Penicilin a streptomycin byly přidány v konečné koncentraci 100 jednotek a 100 ug. Později byla místo Hanks-Simmsova roztoku použita hovězí plodová voda (5). Inhibitor sójového trypsinu v konečné koncentraci 0,05 mg byl přidán do média pro všechny kultury. Do každé kultivační zkumavky obsahující 10 až 15 fragmentů tkáně byl přidán jeden ml média. Kultivační tekutiny se měnily každé 3 až 4 dny. V době inokulace agens bylo médium upraveno tak, aby obsahovalo 85 % Hanks-Simmsovy nebo hovězí amniové tekutiny, 10 % extraktu hovězího embrya a 5 % koňského séra.
B. Stacionární kultury opičích ledvin byly připraveny trypsinizační technikou podle Youngnera(6) s hovězí amniovou tekutinou obsahující 5 % extraktu z hovězího embrya a 10 % koňského séra. Po standardizaci buněčných suspenzí(7) bylo do kultivačních zkumavek přidáno 0,5 ml obsahujících přibližně 500 000 buněk. Po dobrém růstu buněk (7 až 9 dní) bylo médium změněno na 95 % hovězí amniové tekutiny a 5 % koňského séra.
C. HeLa buněčný kmen nádorových epiteliálních buněk, laskavě dodaný Dr. J. T. Syvertonem z University of Minnesota, byl kultivován podle popisu Scherera, Syvertona a Geye(8) a Syvertona (9). Do kultivačních zkumavek bylo nasazeno přibližně 40 000 buněk do 1 ml a za 4 až 5 dní byla získána vrstva růstu obsahující 90 000 až 160 000 buněk. Poté byla živná tekutina nahrazena udržovacím roztokem obsahujícím 10% kuřecí sérum(8,lO). Při delší inkubaci kultur v této tekutině byly zaznamenány degenerativní změny (granulace, zaoblení buněk). Když k tomu došlo, bylo udržovací médium nahrazeno živnou tekutinou, která obnovila normální morfologii za 1 až 2 dny. Proto byla při pokusech s dlouhými inkubačními dobami živná tekutina nahrazena udržovacím médiem a ponechána na buňkách po krátkou dobu.
“Obnova látek MK1, MK3 a MK4 z neinokulovaných kultur opičích ledvin. Krátce po setkání s agens MK-D při pokusech o adaptaci viru dengue na kultury opičích ledvin byly v neinokulované kultuře válečkových zkumavek 12 dní po její přípravě opět pozorovány syncytiální hmoty a vakuoláty.
Tato kultura a dalších 16 kultur připravených ze stejné ledviny byly vyčleněny pro další studium. Patnáctý den byla v této kultuře patrná degenerace, která se objevila i ve třech dalších kulturách, a 34. den v sedmi z 16 kultur. Tekutiny byly odebrány 14. a 15. den z kultur vykazujících degeneraci, byly sloučeny a 0,2 ml bylo inokulováno do buněčných kultur HeLa. Lytické rameno se objevilo za 10 dní. Tekutiny z těchto kultur byly poté odebrány, sloučeny a 0,1 ml přeneseno do čerstvých kultur HeLa. K podobné degeneraci při tomto druhém průchodu došlo za 6 dní. Následné pasáže v buňkách HeLa byly provedeny s 0,1 až 0,2 ml sdružených tekutin odebraných v intervalech 1 až 2 dnů po dobu 4 až 6 dnů po definitivních známkách degenerace. Tekutiny byly uchovávány v COz skříni po různě dlouhou dobu mezi pasážemi. Tento prostředek, označený MK1, má
byl nyní 101krát sériově pasážován v buněčných kulturách HeLa. Agens MK3 a MK4 byla podobně získána z neinokulovaných kultur připravených z ledvin různých opic rhesus. MK3 byl tedy získán ze série neočkovaných kultur, v nichž 8 ze 72 válečkových zkumavek vykazovalo střední až silnou degeneraci 12 dní po přípravě, a MK4 z 11 ze 41 stacionárních kultur s degenerací 20 dní po přípravě s trypsinizovanou buněčnou suspenzí. Tyto 2 látky byly nyní v kulturách HeLa sériově podány 10krát, resp. 6krát. MK3 po 10 průchodech HeLa buněk způsobil vakuolizaci a tvorbu syncytiálních hmot v kulturách opičích ledvin. Stejných výsledků bylo dosaženo s MK4 po 6 pasážích buněk HeLa. Byly získány informace, které naznačují, že četnost výskytu těchto látek v neinokulovaných kulturách může být poměrně vysoká. Tak v 6 z 9 sérií kultur připravených z ledvin různých opic vykazovala jedna nebo více ze 7 až 41 kultur v každé sérii držených 20 až 55 dní charakteristickou degeneraci (tabulka I); a obnovení agens z kultur 3 různých sérií bylo úspěšné. Jeden pokus o obnovu činidla z neočkovaných kultur série ledvin, které nevykazovaly degeneraci, byl neúspěšný po 3 zaslepených pasážích v buňkách HeLa a následné pasáži v kulturách opičích ledvin. Prakticky ve všech kulturách připravených z různých opičích ledvin jsme důsledně pozorovali občasné malé vakuoly patrné od počátku vyrůstání buněk. V některých kulturách přetrvávaly, ale v jiných zřejmě při další inkubaci zmizely.”
“Tato zjištění poskytují důkaz, že tyto látky byly získány z ledvinové tkáně opic. Důkaz, že zdrojem agens nejsou jiné složky tkáňových kultur, je následující: Jedno agens bylo obnoveno v době, kdy byl místo hovězí amniové tekutiny použit Hanksův-Simmsův roztok, a u dalších 3 agens tomu bylo naopak. Extrakt z hovězího embrya byl rutinně používán pro kultivaci HeLa buněk po dobu více než jednoho roku, aniž by došlo k progresivní charakteristické degeneraci, která nastala po inokulaci těchto kultur agens. Koňské sérum bylo před použitím jako složka média zahříváno po dobu 30 minut na 56 °C a, jak již bylo uvedeno, 2 ze 4 takto ošetřených činidel nevyvolaly po 21 dnech cytopatické účinky. Konečně 3 slepé pasáže v HeLa buňkách a následná pasáž v opičích ledvinových kulturách tekutin z jedné série opičích ledvin neodhalily agens. Agens však bylo získáno z kultur dvou dalších sérií opičích ledvin, které byly vytvořeny krátce před a po této sérii se stejnými šaržemi složek média.”
“Shrnutí. Při pokusech o adaptaci viru dengue na kultury opičích ledvin rhesus bylo zjištěno neidentifikované agens, které v těchto kulturách způsobuje tvorbu syncytiálních hmot a vakuolizaci. Následně byly z neinokulovaných kultur opičích ledvin předány a udržovány v kulturách HeLa buněk 3 další agens s podobnými cytopatogenními účinky. Zdrojem agens je ledvinová tkáň, nikoliv složky média. Bakteriologické studie s jedním agens byly negativní. Tentýž agens prošel Selasovým filtrem. V souladu s tím se má za to, že má virovou povahu. Podobné pokusy nebyly provedeny s dalšími 3 agens, ale vzhledem k určitým společným vlastnostem se předpokládá, že jsou stejné povahy.”
rustigian1955 Stáhnout

V této druhé studii, rovněž z roku 1955, Cohen a kol. zjistili u svých neinokulovaných kontrolních kultur stejný cytopatogenní účinek, jaký byl pozorován u jejich inokulovaných vzorků údajně obsahujících “virus” spalniček. To opět ukázalo, že CPE nebyl specifickým účinkem způsobeným nějakým “virem”, a byl tedy vyvolán experimentálními podmínkami. Cohen a spol. si uvědomili, že tento efekt nemohou použít k rozpoznání spalničkového “viru”, a proto tvrdili, že výsledky protilátek, o nichž předpokládali, že jsou způsobeny spalničkovým “virem”, jsou dostatečným kritériem pro rozlišení kultivovaných vzorků:
Fluorescenční protilátky a testy fixace komplementu u agens izolovaných v tkáňových kulturách pacientů se spalničkami.
“Enders a Peebies(1) pozorovali, že tkáňové kultury lidských nebo opičích ledvin inokulované vzorky od pacientů se spalničkami podléhají po jedné nebo více pasážích charakteristickým jaderným a jiným cytologickým změnám. V kulturách se současně objevil jak infekční virus, tak specifický antigen fixující komplement a cytopatogenní účinek byl neutralizován spalničkovým sérem v rekonvalescentní fázi. Na podobné kultury jsme použili techniku fluorescenčních protilátek Coonse a jeho kolegů (2,3) a zjistili jsme, že výsledky jsou paralelní s výsledky komplement-fixačních testů, a poskytují tak imunochemickou podporu pro údaje Enderse a Peeblese a navíc důkaz, že spalničkový (resp. spalničkové) antigen (antigeny) jsou přítomny v jádrech i v cytoplazmě infikovaných buněk.
Metody. Pacienti byli vybráni ke studiu během mírné epidemie typických zarděnek. Klinická diagnóza byla v některých případech podpořena rozpoznáním obrovských buněk Warthinova-Finkeldeyova typu na barvených snímcích nosního výtoku získaných na počátku onemocnění, jak popsali Tompkins a Mcaulay(4).
Odběr vzorků a příprava tkáňových kultur. Byly použity postupy popsané Endersem a Peeblesem (1). Krev, výtěry z krku nebo výplachy nebo nosní výtok byly získány před výskytem vyrážky nebo do jednoho dne po něm. Pokud to bylo možné, byly vzorky inokulovány do tkáňových kultur během několika hodin po jejich odběru. Tkáňové kultury se skládaly z trypsinizovaných buněk opičích ledvin pěstovaných ve fóliích (5). Do zkumavek se inokulovalo 0,2 nebo 0,3 ml připravených vzorků, doplnilo se živnou tekutinou, aby objem dosáhl 1,0 ml, a inkubovalo se jako stacionární preparáty při 36,5 °C. Živná tekutina se skládala z hovězí plodové vody obsahující 5 % inaktivovaného koňského séra a 5 % extraktu z hovězího embrya. První pasáž obsahovala 100 jednotek penicilinu, 100 ug streptomycinu a 50 ug nystatinu (fungicidinu) (6): ostatní 50 jednotek a 50 a 10 ug. Převody do čerstvých zkumavek byly prováděny ve 14denních intervalech s 0,2 ml směsných tekutin odebraných 5, 9 a 14 dní po inokulaci. Kontroly se skládaly z neočkovaných kultur a z kultur, které byly sériově pasážovány s tekutinami z neočkovaných zkumavek.
Výsledky. Přenosná agens, pravděpodobně viry, byla izolována jak z krve, tak z výtěru z krku jednoho pacienta a z výtoku z nosu a výtěru z krku dalších dvou pacientů, jejichž krev nebyla získána. Vzorky výtěrů z krku byly uchovávány v ledové komoře po dobu 7 dnů. Ve vzorcích z dalších 5 případů nebyly zjištěny žádné agens. Inokuly od 3 z nich byly před kultivací uchovávány při teplotě 4°-6 °C po dobu 2 dnů; testy inokul od dalších 2 byly přerušeny po 3 zjevně negativních sériových pasážích. Tyto okolnosti mohly přispět k tomu, že se nám nepodařilo izolovat původce.
Enders a Peebles(1) a Rustigian et al. (10) se setkali s latentními virovými agens, která vyvolávají výraznou vakuolizaci a syncytiální hmoty v tkáňových kulturách opičích ledvin. Buněčná degenerace charakteristická pro tyto “opičí ledvinové agens” se často objevovala v našich kulturách a to jak v těch, které byly očkovány vzorky od pacientů se spalničkami, tak v kontrolních kulturách; proto bylo obtížné použít cytologická kritéria pro rozpoznání spalničkových agens. V některých sériích tkáňových kultur “monkey-kidney agens” zničili buněčné vrstvy za 10 až 14 dní. Při identifikaci 3 původců kultivovaných z inokula spalniček jsme se proto spolehli na testy na přítomnost spalničkového antigenu.”
“Diskuze. Prokázání, že tkáňové kultury inokulované vzorky od pacientů se spalničkami produkují antigeny, které specificky reagují se spalničkovými séry v rekonvalescentní fázi, dokládá zjištění Enderse a Peeblese( 1). Jejich filtrační pokusy jsme neopakovali, ale nepodařilo se nám z infikovaných tkáňových kultur získat plísně, uzliny z králíků imunizovaných záškrtem ani bakterie včetně organismů podobných pleuropneumonii a leptospiry a proto předpokládáme, že antigeny, které vyvolaly specifické reakce ve fluorescenčních testech na protilátky a komplement-fixaci, pocházejí z viru spalniček. Celkově byl antigen v infikovaných tkáňových kulturách detekován dříve metodou fluorescenčních protilátek než komplement-fixačními testy. Předběžné údaje však naznačují, že tyto testy mají význam v diferenciální diagnostice exantémů a encefalitid neznámého původu.”
cohen1955 Stáhnout
Tato poslední studie, na kterou Dr. Cowan upozornil, pochází od Von Magnuse a kol. z roku 1959. V souladu s předchozími výzkumníky narazili na naprosto stejný cytopatogenní účinek i u neočkovaných kultur ze zdravých opic. Výzkumníci tvrdili, že změny byly způsobeny něčím, co označili jako “pěnotvorné činidlo”, a uvedli, že tato “činidla” se pravidelně vyskytují ve zdravých opičích ledvinových buňkách. Vzniklý CPE byl k nerozeznání od toho, který byl spojen s “virem” spalniček. Von Magnus a spol. se tedy stejně jako před nimi Cohen a spol. rozhodli, že tento efekt má při pokusu o identifikaci “virů” omezený význam, a místo toho se při určování, zda je ve vzorku přítomen “virus” spalniček, spolehli na nespolehlivé testy na protilátky. Zajímavé také je, že tito výzkumníci selhali při četných pokusech o “izolaci” spalničkového “viru”, neboť byli “úspěšní” pouze v 5 ze 13 případů. Všechny pokusy o izolaci “viru” selhaly 24 hodin po nástupu vyrážky. Bez ohledu na své neúspěchy se Von Magnus a spol. rozhodli, že jejich nepřímé důkazy souhlasí s předpokladem, že výsledky vyšetření protilátek identifikovaly “virus” spalniček: “Spalničky” byly identifikovány na základě výsledků vyšetření protilátek.
Studie viru spalniček v tkáňových kulturách opičích ledvin. 1. Izolace viru od 5 pacientů se spalničkami.
“Jak popsali Enders & Peebles (6) a později Rustigian et al. (13) a Cohen el al. (3), cytopatické změny podobné těm, které způsobuje virus spalniček, lze pozorovat i v neinokulovaných kulturách tkáně opičích ledvin (obr. 4-5). Tyto změny jsou pravděpodobně způsobeny virům podobnými agens, tzv. pěnovými agens, které se zřejmě často vyskytují v ledvinových buňkách zdánlivě zdravých opic. Specifický spalničkový antigen je však produkován pouze v kulturách infikovaných virem spalniček. V této studii byla proto jako kritérium přítomnosti spalničkového viru použita schopnost materiálu z pasáží tkáňových kultur fixovat komplement v přítomnosti spalničkového séra v konvalescentní fázi.”
MATERIÁLY A METODY
Tkáňové kultury: Běžně byly použity buňky trypsinizovaných ledvin opic rhesus, opic cynomolgous nebo paviánů. Pro přípravu nebo těchto tkáňových kultur byla použita technika Youngerova (15) s některými modifikacemi, jak bylo dříve popsáno (10). Během růstu buněk se růstové médium skládalo z 0,5 % hydrolyzátu laktalbuminu (11) v Hankově roztoku (5) s 2 % koňského séra.
Před výsevem viru bylo médium vyměněno. Každá zkumavka obdržela 1,8 ml syntetického média 199 (12) nebo hovězí amniové tekutiny (5) obsahující fenolovou červeň jako indikátor (konečné ředění 0,02 %). Všechna média obsahovala penicilin (100 U/ml) a streptomycin (0,1 mg/ml).
Odběr vzorků: pokus o izolaci viru byl proveden z výplachů hrdla a krve.
Výplachy z hrdla: Pacienti se spalničkami byli požádáni, aby kloktali buď 15 ml směsi jednoho dílu infuzního bujónu z volského srdce a dvou dílů pufrovaného solného roztoku, nebo v pozdějších pokusech 15 ml destilované vody obsahující l % Bacto tryptózy “Difco”. Do tekutiny byl přidán penicilin 100 U na ml a streptomycin 0,1 mg na m l. Z krku velmi malých dětí byly získány vzorky pomocí vatových tamponů, které byly následně ponořeny do 2 ml jedné z obou právě popsaných tekutin. Ve všech případech byly vzorky okamžitě zmraženy v C02-ledku a poté uloženy do elektrického mrazáku (-60 °C). Před inokulací do tkáňové kultury byl materiál rychle rozmrazen při 37 °C v tekoucí vodě z vodovodu.
V dřívějších pokusech byl do krve přidán heparin (2 ml O.O5 % heparinu na 10 ml krve). Při pozdějších pokusech se krev nechala srazit. Červené krvinky, které se nesrazily, byly resuspendovány v séru a tato směs byla použita jako inokulum pro tkáňové kultury. Vzorky krve se uchovávaly při teplotě +4 °C až do doby inokulace.
Inokulace tkáňových kultur: Tkáňové kultury byly inokulovány 0,5 rnl materiálu z výplachu hrdla nebo 0,25 ml krve. Konečné množství tekutiny ve zkumavkách bylo asi 2 ml. Kultury byly uchovávány ve stacionární poloze nebo byly umístěny do rotujícího bubnu (1 otáčka za minutu).
Subkultury z první pasáže se prováděly mezi 6. a 16. dnem, obvykle 8. den po inokulaci. Živné médium se v průběhu pasáže zpravidla nevyměňovalo. Subkultury z pozdějších pasáží se prováděly mezi 6. a 12. dnem inkubace. Materiál z pasáže se skládal ze suspenze buněk a buněčných zbytků v kultivačním médiu. Tato směs byla získána uvolněním buněk dosud přiléhajících ke sklu seškrábnutím pipetou. Množství inokulované směsi se lišilo, ale obvykle činilo 0,2 mi, takže konečný objem tekutiny v inokulovaných zkumavkách byl asi 2 ml. Stejným způsobem se prováděly sériové pasáže materiálu z kontrolních zkumavek.
DISKUSE
“Tato práce potvrzuje zjištění Endel’s & Peebles (G) a Cohena et al. že výplachy hrdla a krevní výpotky pacientů v raném stadiu spalniček obsahují virus, který je schopen vyvolat charakteristické cytopatické změny v kulturách lidských a opičích ledvinových buněk. V inokulovaných kulturách byly pozorovány stejné syncyklické útvary, jaké popsali Enders & Peebles (6). Kromě toho bylo zjištěno, že v systému tkáňových kultur použitém v této práci se při pokračující inkubaci vyvinuly další specifické změny. Tento druhý krok degenerace vedl k nahromadění buněčných zbytků s kruhovými zrnitými útvary, které měly buď zcela hladký, nebo zvrásněný okraj. Zdá se, že tyto cytopatické změny jsou pro virus spalniček stejně specifické jako synchronie.
Nicméně opičí ledvinové viry nebo “pěnivé agens” mohou vyvolat buněčné degenerace, které jsou mikroskopicky k nerozeznání od degenerací způsobených virem spalniček. Z tohoto důvodu mají cytologické projevy při studiu spalniček omezenou hodnotu a ke stanovení identity kultivovaných původců jsou zapotřebí další kritéria. Toho lze dosáhnout průkazem intranukleárních inkluzních tělísek v buněčných kulturách (6) nebo testy na přítomnost spalničkového antigenu pomocí techniky fluorescenčních protilátek (3) nebo reakce fixace komplementu (3, 6). Poslední zmíněná metoda byla pro svou jednoduchost použita v této studii. Pozorování vývoje spalničkového antigenu v různých kulturách budou podrobně popsána v připravované publikaci (1).
Ve studii prezentované v tomto článku byl virus spalniček izolován z pěti z devíti výplachů hrdla odebraných 24 hodin po propuknutí exantému. Důvodem, proč se izolace nepodařila u jednoho pacienta (č. 12), může být to, že bezprostředně před kloktáním zvracel. Ve zbývajících třech případech (č. 2, 7 a 9) nelze pro negativní výsledky nabídnout žádné zjevné vysvětlení. Je možné, že to bylo proto, že všechny tyto izolace byly provedeny pomocí výtěrů z krku u malých dětí, které byly velmi rozrušené, a odběr vzorků se tak obtížně prováděl.
Virus byl z krve získán pouze v jednom z osmi pokusů provedených do 24 hodin po výskytu vyrážky. Tato míra izolace je nízká ve srovnání s mírou izolace, kterou získali Enders & Peebles (6), kteří získali virus z krve u čtyř z pěti pacientů. Tito autoři použili heparinizovanou krev, ze které je možná snazší získat virus než ze séra obsahujícího resuspendované krevní buňky. V naší laboratoři byla jediná izolace z krve provedena z jednoho ze tří vzorků, do kterého byl přidán heparin. Enders a Peebles také použili větší inokulum (0,5 ml až 2 ml) než v této studii (0,25 ml).
Zvažovala se možnost, že neúspěch při získávání viru od 8 ze 13 zkoumaných pacientů může být způsoben necitlivostí konkrétních použitých buněk vůči viru spalniček. To však zřejmě nebyl tento případ, protože když byly zkumavky připravené současně se zkumavkami použitými v neúspěšných izolačních pokusech inokulovány tekutinou infikovanou spalničkami, objevily se typické cytopatické změny spolu s výskytem antigenu vázajícího komplement v živném médiu.
Pozorování uvedená v tomto článku dobře souhlasí s předpokladem, že izolovaná agens jsou příčinou spalniček.”
SHRNUTÍ
(1) Viroví agens byli izolováni v trypsinizovaných kulturách tkáně opičích ledvin z výplachů hrdla a krve 5 ze 13 pacientů vyšetřených během akutní fáze spalniček. Ve všech případech byl virus izolován z výplachů hrdla nebo krčních výplachů, zatímco jeden kmen byl získán z krve. Všechny pokusy o izolaci viru později než 24 hodin po nástupu vyrážky selhaly.
2) Jsou popsány cytopatické projevy pozorované u tkáňových kultur infikovaných virem spalniček i u neinokulovaných zkumavek. Jako kritérium přítomnosti tohoto viru v infikovaných kulturách byly použity testy fixace komplementu na přítomnost spalničkového antigenu.
(3) Intranazální a orální podání materiálu z pozdních pasáží jednoho z izolovaných původců dvěma rhesusům vedlo u jednoho ze zvířat k výrazné vyrážce podobné spalničkám a u obou opic se vyvinuly protilátky proti inokulovanému kmeni.
(4) V sérologických studiích sér pacientů s akutní a rekonvalescentní fází spalniček, při nichž byly jako antigen v komplement fixačních testech použity kultury opičí ledvinové tkáně infikované virem spalniček, byl ve všech případech pozorován zřetelný vzestup protilátek.
bech-von-magnus-1959-1 Stáhnout

Identické CPE u neočkovaných (vlevo) a očkovaných (vpravo) vzorků.
Z těchto tří studií v kombinaci s Endersovými vlastními neočkovanými kontrolami si můžeme odnést, že k tomu, aby se projevil cytopatogenní účinek, nebyl nikdy nutný žádný “virus”. Tento účinek byl pozorován v kulturách ledvinových buněk zdravých opic. Bylo konstatováno, že tyto nálezy nejsou vzácné, protože pozorování tohoto cytopatogenního účinku v neočkovaných kulturách ze zdravých opic bylo ve skutečnosti poměrně časté. Je tedy zřejmé, že k odumření buněk vede samotná metoda kultivace, která spočívá ve stresování a hladovění buněk, které se nacházejí mimo své přirozené prostředí, a jejich prolévání ledvinově toxickými antibiotiky, jakož i přimíchávání cizorodých živočišných materiálů a dalších chemických přísad. Vzhledem k tomu, že tento cytopatogenní účinek nebyl specifický pro “virus” a objevil se v kulturách bez přítomnosti jakýchkoli “virů”, nelze jej použít k tvrzení o přítomnosti “viru”, jak uvedli Cohen et al. i Von Magnus et al. Žádné výsledky získané z tohoto pseudovědeckého experimentu tedy nemají žádnou hodnotu.

Ačkoli by výše uvedené studie měly stačit k tomu, aby smyly veškeré pochybnosti, které zůstaly o podvodném používání cytopatogenního efektu jako potvrzení přítomnosti “viru” ve vzorku, přihazuji do placu také publikaci Hulla a kol. z roku 1956, která poskytuje ještě více usvědčující důkazy podporující ukončení této pseudovědecké praxe. V této práci vědci uvádějí, že používání metody buněčných kultur vedlo k objevu mnoha neznámých cytopatogenních agens, která lze zjistit pouze pomocí tohoto postupu. Výzkumníci použili drobné odchylky v CPE pozorované během kultivace vakcín proti poliomyelitidě k tvrzení, že v kulturách bylo v různých obdobích přítomno nejméně 10 dalších neviditelných “virům podobných agens”. Ačkoli se Hull a spol. domnívali, že tyto agens s největší pravděpodobností pocházejí z buněk opičích ledvin, uvedli, že existuje možnost, že tyto “agens” mohou představovat kontaminanty z lidských zdrojů, koňského séra, živného média nebo jiných roztoků použitých při přípravě kultur. Bez ohledu na to rozhodli, že tito neviditelní agens, kteří byli “získáni” z normálních nebo neinokulovaných kontrolních kultur zdravých opic, jsou “opičí viry”. Výzkumníci tak učinili navzdory skutečnosti, že se jim nepodařilo různými způsoby injekčního podání vyvolat u žádného zvířete onemocnění až na dvě výjimky, kdy dva z “virů” způsobily po injekci do mozku smrt. Po injekci těchto dvou “virů” do svalů však k žádnému onemocnění nedošlo. Jinými slovy, Hull a spol. pozorovali cytopatogenní účinky, které subjektivně určili, že nejsou způsobeny “virem” obrny jak v inokulovaných, tak v neinokulovaných kulturách a na základě pozorovaného vzoru buněčné smrti rozhodli, že v kultuře musí být přítomny různé “viry”. U těchto směsí, u nichž se předpokládalo, že obsahují neviditelné “viry”, se důsledně prokázalo, že nejsou patogenní a proto nepozorovatelné entity z neinokulovaných kultur se stejným obrazem CPE ze zdravých opic nesplňovaly definici “viru”:
Virové agens získané z tkáňových kultur opičích ledvinových buněk
“Zvýšené používání techniky kultivace buněk pro izolaci, udržování a studium virů vedlo k objevu mnoha dosud neznámých cytopatogenních agens. Při screeningu vzorků lidské stolice nebo rektálních výtěrů na viry poliomyelitidy izoloval Melnick (1) několik “osiřelých virů”, které se netypovaly s antiséry poliovirů, a ze stejného zdroje získal Sabin (2) 5 virů označovaných jako He1; He2 atd. Novou řadu virů RI izolovali na tkáňových kulturách Hilleman a Werner (3) z výplachů hrdla pacientů s respiračním onemocněním. Rowe, Huebner a další (4) izolovali podobnou skupinu virů, skupinu APC, které se nejprve objevily v tkáňových kulturách lidské adenoidní a tonzilární tkáně a později byly získány z výplachů očí pacientů se zánětem spojivek. Nedávno Rustigian et al. (5) oznámili obnovení dvou cytopatogenních agens z kultur buněk opičích ledvin. Ve tkáních a výkalech člověka a nižších živočichů je tedy přítomna řada virových agens, která se brání detekci jinými metodami než tkáňovými kulturami.
Během výroby a testování vakcíny proti poliomyelitidě byly v našich laboratořích a v dalších laboratořích účastnících se tohoto programu pozorovány statisíce kultur opičích ledvin připravených z tisíců opic. Vyskytla se také četná filtrovatelná, přenosná cytopatogenní agens jiná než poliovirus. Ačkoli tato agens byla do kultur pravděpodobně vnesena s buňkami opičích ledvin, zůstává vzdálená možnost, že některá z nich představují kontaminanty z lidských zdrojů, koňského séra, živného média nebo jiných roztoků používaných při přípravě kultur. Tato zpráva popisuje 8 imunologicky odlišných původců izolovaných a studovaných v našich laboratořích a odkazuje na 2 původce z jiných zdrojů.”
“Izolovaní původci budou označováni jako “similské viry” (S.V.), dokud nebude možné určit jejich spojení s nějakým jiným hostitelem nebo identifikaci. Tito původci budou označováni jako S.V.1, S.V.2 atd. Z původní skupiny původců, která zahrnovala S.V. až S.V.15, bude v tomto článku popsáno osm (S.V. 1, 2 4, 5. 6, 11, 12 a 15). Chybějící čísla si podržely látky, které byly identifikovány nebo reklasifikovány po studiích antisér, nebo které ještě nebyly dostatečně prozkoumány pro zařazení v této době. Do posledně jmenované kategorie patří S.V.13, který je “krajkový nebo pěnivý” agens, který se často vyskytuje v kulturách opičích ledvinových buněk.
VÝSLEDKY
Následuje stručný popis původní izolace a popisu 8 výše uvedených činidel. S.V.1 byl získán ze zkumavkových kultur trypsinizovaných buněk opičích ledvin, které byly vysazeny 8. února 1954. Tato série zahrnovala 1 700 zkumavek připravených ze souboru ledvin 4 zjevně zdravých opic rodu Cynomolgus. Po 6 až 8 dnech inkubace se kultury jevily jako uspokojivé a byly použity k titraci polioviru. Po dalších 6 dnech inkubace byl v některých kulturách pozorován typ destrukce buněk za koncovým bodem polioviru, který byl netypický pro poškození způsobené poliovirem. Toto atypické poškození bylo pozorováno u 17 % kultur. Tento cytopatogenní účinek (C.P.E.) byl charakterizován zmenšením velikosti a výrazným zaoblením buněk.”
“Opice byly všemi viry inokulovány intracerebrálně, intramuskulárně a v některých případech i jinými cestami. Sérum každé inokulované opice bylo nejprve vyšetřeno na přítomnost protilátek proti specifickému viru, který měl být injektován. Použita byla pouze negativní zvířata. S výjimkou S.V.12 a S.V.15 nebyly pozorovány žádné známky klinického onemocnění nebo hrubých či histopatologických změn. Opice inokulované intracerebrálně neředěnými viry S.V.12 a S.V.15 ve vysokém titru podlehly během 4 až 6 dnů po inokulaci. Specifický typ inokulovaného viru byl snadno získán z mozkových a míšních tkání těchto zvířat. Stejné dva viry však při intramuskulární inokulaci opicím nevyvolaly klinické onemocnění ani hrubé či histopatologické změny. Histopatologické změny pozorované u opic inokulovaných intracerebrálně odhalily nekrózu a úplnou destrukci cévnatého plexu a generalizovaný aseptický typ meningitidy. Nebyla pozorována žádná jiná mozková patologie ani léze připomínající poliomyelitidu. Různé studované orgány nevykazovaly žádné rozpoznatelné léze a zdálo se, že veškeré postižení bylo omezeno na centrální nervový systém v oblasti inokulace viru. Léze způsobené S.V.12 a S.V.I5 byly nerozlišitelné.
Přestože u opic naočkovaných těmito viry nevzniklo žádné rozpoznatelné onemocnění, až na výše uvedené výjimky byly v našich laboratořích i v jiných laboratořích získány určité důkazy o tom, že S.V.2 a S.V6 mohou mít určitý etiologický význam při střevních infekcích.”
“Žádný z těchto opičích virů neinfikoval embryonální vajíčka, jak bylo zjištěno na základě úhynu embrya. Nicméně jeden původce z A.M.S.G.S. identifikovaný jako S.V. byl získán z embryonálních vajíček inokulovaných tkáňovými kulturami opičích ledvin. Skupiny dospělých a sajících myší byly inokulovány každým virem intracerebrálně a intraperitoneálně. Nebyly však pozorovány žádné známky onemocnění ani úhynu myší. Potkani a králíci, kteří byli inokulováni živým virem za účelem výroby antiséra, rovněž nepodlehli infekci žádným z těchto agens.”
DISKUSE
Byly popsány izolace a charakteristiky 8 zřejmě nových virů. Ačkoli nebyl předložen jednoznačný důkaz, je pravděpodobné, že tato agens jsou opičího původu. Jaká onemocnění, pokud vůbec nějaká, by mohly u opic způsobovat, rovněž nebylo zjištěno. Viry S.V.2 a S.V. 6 mohou být etiologickými původci střevních infekcí opic, protože oba byly izolovány ze vzorků stolice zvířat trpících průjmem. Žádný z nich nezpůsobil experimentální infekci při intramuskulární inokulaci, ačkoli S.V.12 i S.V.15 způsobily u opic po intracerebrální inokulaci protrahovaný typ paralýzy a smrt. Pokusy o infikování opic jinými způsoby inokulace nejsou dosud ukončeny.”
Největší význam těchto virů byl jejich výskyt v tkáňových kulturách používaných k výrobě a testování vakcíny proti poliomyelitidě. To bylo zvláště nepříjemné při testování bezpečnosti vakcíny na tkáňových kulturách, protože výskyt těchto virů v mnoha případech znehodnotil testy na poliovirus, což si vyžádalo opakování testů, a tím zpozdilo uvolnění vakcíny. V některých případech se také vyskytl problém s prokázáním, že kontaminující “simian virus” izolovaný během testu bezpečnosti vakcíny nebyl v testovaném vzorku vakcíny. Obecně to však nebyl problém, protože mnoho těchto původců bylo získáno z normálních nebo neočkovaných kontrolních kultur.”
SHRNUTÍ
“Bylo popsáno osm zřejmě nových submikroskopických, filtrovatelných, cytopatogenních agens získaných v tkáňových kulturách ledvinové tkáně opic rhesus a cynomolgus. Mezi žádným z těchto agens a přirozeně se vyskytujícími chorobami opic nebo jiných zvířat nebyla zjištěna žádná jednoznačná souvislost. Byl diskutován význam těchto virů v programu vakcinace proti poliomyelitidě a problémy, které způsobují při testování bezpečnosti vakcín na tkáňových kulturách.”
hull1956 Stáhnout
Tyto studie by samy o sobě měly stačit k tomu, aby ukázaly, že při kontrolách se ukáže, že pozorovaný cytopatogenní účinek, který je připisován přítomnosti neviditelného “viru”, není specifický pro žádný “virus”. Ve skutečnosti nejsou k vyvolání tohoto účinku vůbec žádné “viry” zapotřebí. Naštěstí máme to štěstí, že se Dr. Stefan Lanka při prokazování této skutečnosti nespoléhal jen na práci jiných a vzal si za své ověřit tato zjištění provedením vlastní sady kontrolních pokusů, čímž zatloukl pověstný poslední hřebíček do rakve této pseudovědecké praxe. Protože Dr. Cowan odvedl naprosto vynikající práci při rozebírání kontrolních experimentů Dr. Lanky, uvádím výňatky z jeho knihy Breaking the Spell (CZ verze ZDE):

“Zde je podstata Lankova experimentu, provedeného nezávislou profesionální laboratoří, která se specializuje na kultivaci buněk. Jak je vidět na této sérii fotografií, každý ze čtyř svislých sloupců je samostatný experiment. Horní fotografie v každém sloupci byla pořízena první den a spodní fotografie byla pořízena pátý den.
Ve svislém sloupci jedna byly normální buňky kultivovány s normálním živným médiem a pouze malým množstvím antibiotik. Jak vidíte, ani první, ani pátý den nebyl zjištěn žádný CPE; buňky pokračovaly v normálním, zdravém růstu.
Ve svislém sloupci dva byly normální buňky opět pěstovány na normálním živném médiu a malém množství antibiotik, ale tentokrát bylo k obohacení média přidáno 10% fetální telecí sérum. Přesto buňky v kultuře rostly normálně, a to jak první, tak pátý den.
Třetí svislý sloupec ukazuje, co se stalo, když skupina doktora Lanky použila stejné postupy, které byly použity při každém moderním experimentu s izolací každého patogenního viru, který jsem viděl. To zahrnovalo změnu živného média na “minimální živné médium” – což znamená snížení procenta fetálního telecího séra z obvyklých 10 % na 1 %, což snižuje množství živin dostupných pro růst buněk, a tím je stresuje – a ztrojnásobení koncentrace antibiotik. Jak vidíte, pátý den experimentu se objevil charakteristický CPE, který “dokazuje” existenci a patogenitu viru – až na to, že v žádném okamžiku nebyl do kultury přidán patogenní virus. Tento výsledek může znamenat pouze to, že CPE byl důsledkem způsobu provedení kultivačního pokusu a nikoliv nějakého viru.
Čtvrtý a poslední svislý sloupec je stejný jako svislý sloupec třetí, s tím rozdílem, že k této kultuře byl přidán roztok čisté RNA z kvasinek. To vedlo ke stejnému výsledku jako ve sloupci tři, což opět dokazuje, že příčinou CPE je technika kultivace – a nikoli virus.“
Neplatný experiment = neplatné kontroly

Nyní by snad již mělo být jasné, proč má virologie problém s kontrolami. Samotná metoda buněčné kultivace není platným experimentálním uspořádáním, protože nikdy nebyla navržena v souladu s vědeckou metodou. Experiment vytváří účinek (CPE) a poté předpokládá příčinu (“virus”), aniž by se ověřilo, že předpokládaná příčina vůbec existuje. I kdybychom jim dali volnost při použití buněčné kultury jako platného experimentu k prokázání přítomnosti “viru”, je známo, že cytopatogenní účinek je způsoben mnoha dalšími faktory, které s žádným “virem” nesouvisejí, takže vysvětlení fiktivního “viru” jako viníka je zcela zbytečné. Na obviňování neviditelných “virů” z pozorovaného CPE namísto jiných vysvětlení bylo poukázáno v technickém bulletinu pro kultivaci buněk:
Pochopení a zvládání kontaminace buněčných kultur
“Vzhledem k tomu, že cytopatické viry obvykle ničí kultury, které infikují, mají tendenci se samy omezovat. Proto když se kultury samy zničí bez zjevné příčiny a nelze nalézt žádné důkazy o běžných biologických kontaminantech, jsou často obviňovány kryptické viry. (Viz obrázky 3a a 3b.) Jsou to dokonalí viníci, neviditelní a nezjistitelní; viníci bez přímých důkazů. To je nešťastné, protože skutečnou příčinou zničení této kultury může být něco jiného, možná mykoplazma nebo chemická kontaminující látka, a v důsledku toho zůstane neodhalena, aby se stala závažnějším problémem.“

“Vina bez přímých důkazů.”
Je známo, že CPE mohou způsobovat i jiné faktory než “viry”, např:
- Bakterie
- Améby
- Paraziti
- Antibiotika
- Antimykotika
- Chemické kontaminanty
- Stáří a poškození buněk
- Stres způsobený prostředím
A abych nezapomněl, cytopatogenní účinky prý vytvářejí i smyšlené “virům podobné” entity (o nichž se přiznává, že je nelze oddělit od “virů”) známé jako exozomy.

Cytopatogenita EVs z Acanthamoeba castellanii na buňkách C6. a Krysí gliové buňky C6 byly kokultivovány s EVs z A. castellanii a CPE byl hodnocen barvením podle Giemsy po 24 h. Údaje jsou reprezentativní pro tři nezávislé experimenty. b Světelné mikrofotografie ukazují CPE buněk C6, které byly vytvořeny EVs z A. castellanii po 4 h, 8 h, 12 h, 16 h a 20 h. c Světelné mikrofotografie ukazují CPE buněk C6, které byly vytvořeny EVs z A. castellanii po 4 h, 8 h, 12 h, 16 h a 20 h. d Světelné mikrofotografie ukazují CPE buněk C6, které byly vytvořeny EVs z A. castellanii po 24 h.
Zopakujme, že cytopatogenní účinek není platnou závislou proměnnou, protože se nejedná o přirozeně pozorovaný jev a lze jej vysvětlit různými jinými faktory než neviditelným “virem”. Nepřečištěné tekutiny použité během kultivace nejsou platnou nezávislou proměnnou, protože nikdy nebylo prokázáno, že “virus”, o němž se předpokládá, že se v nich nachází, existuje v přečištěném a izolovaném stavu před provedením pokusu. Provádění buněčné kultury jako důkazu existence “viru” je tedy zcela nevědecké, protože nelze postupovat podle vědecké metody. Nicméně v zájmu argumentace připusťme, aby virologové měli buněčnou kulturu a cytopatogenní účinek jako platný experiment. To by znamenalo, že by museli mít pokaždé vedle buněčných kultur provedeny i platné kontroly. Člověk by si myslel, že to bude běžně k vidění ve virologických pracích, jak jsme to viděli v těch z 50. let minulého století. Častěji se však stává, že v rámci studií předkládaných jako důkaz existence “virů” buď nikdy nenajdeme žádnou zmínku o kontrolách, nebo je špatně definováno, co bylo s kontrolní kulturou provedeno. Možná je to způsobeno katastrofálními závěry autorů studií z 50. let, které nenapravitelně poškodily tvrzení o “virové” příčině pozorovaného CPE? V každém případě, pokud virologové provádějí kontrolu, obvykle provádějí tzv. maketové infekce:
mock-infected
“Kontrola používaná při experimentech s infekcemi. Používají se dva vzorky, z nichž jeden je infikován zájmovým virem/vektorem a s druhým se zachází stejným způsobem, jen bez viru. Někdy se pro mock-infekci použije nevirulentní kmen.”
https://www.genscript.com/biology-glossary/10558/mock-infected
To znamená, že virologové mají použít stejnou kulturu se stejnými přísadami (antibiotika, antimykotika, minimální živná půda, fetální hovězí sérum atd.), ale bez přidaného “viru”. Podívejme se na několik příkladů, abychom zjistili, zda to tak skutečně dělají. V tomto prvním případě autoři studie CDC uvádějí, že mock-infikovaná kultura byla ošetřena pouze médiem, takže se zdá, že to odpovídá výše uvedené definici. Neuvádí se však, zda bylo médium přesně stejné jako u experimentální kultury. Musíme předpokládat, že tomu tak je, což je problém, jak bude ukázáno později.
Replikace koronaviru spojeného se SARS v buněčných liniích

https://wwwnc.cdc.gov/eid/article/12/1/05-0496_article
Tato druhá studie je jednou z klíčových raných studií “SARS-COV-2” od Zhu et al. Dozvídáme se pouze, že kontrola je maketová infekce, a proto musíme opět předpokládat, že výzkumníci použili přesně stejné médium/složky, protože žádné další podrobnosti nebyly uvedeny.
Nový koronavirus od pacientů s pneumonií v Číně
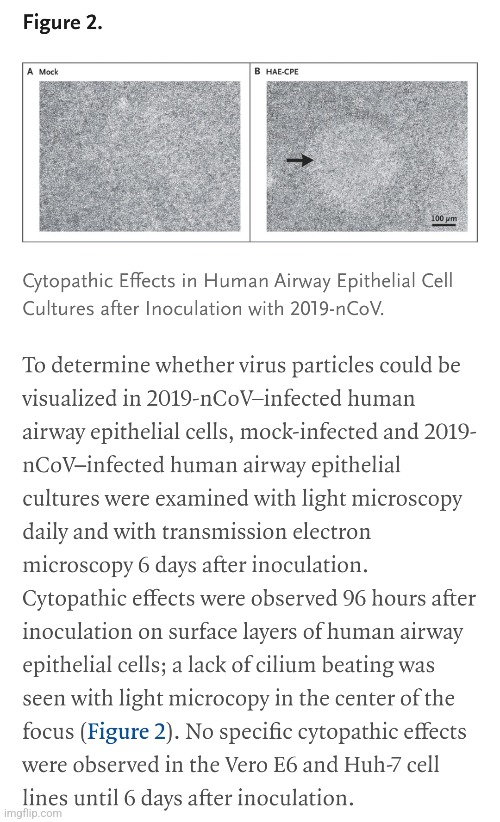
https://www.nejm.org/doi/full/10.1056/nejmoa2001017
Třetí příklad pochází z australské studie z dubna 2020, která je nechvalně proslulá tím, že do kultury přidala trypsin, proteinový trávicí enzym, aby se na EM snímcích vytvořila “hrotovitá” korona. Výzkumníci nám poskytují tzv. neinfikované kontrolní buňky. Opět musíme předpokládat, že s neinfikovanou buňkou bylo zacházeno úplně stejně jako s infikovanou buňkou, protože nebyly uvedeny žádné podrobnosti.
Izolace a rychlé sdílení nového koronaviru 2019 (SARS-CoV-2) od prvního pacienta s diagnózou COVID-19 v Austrálii.
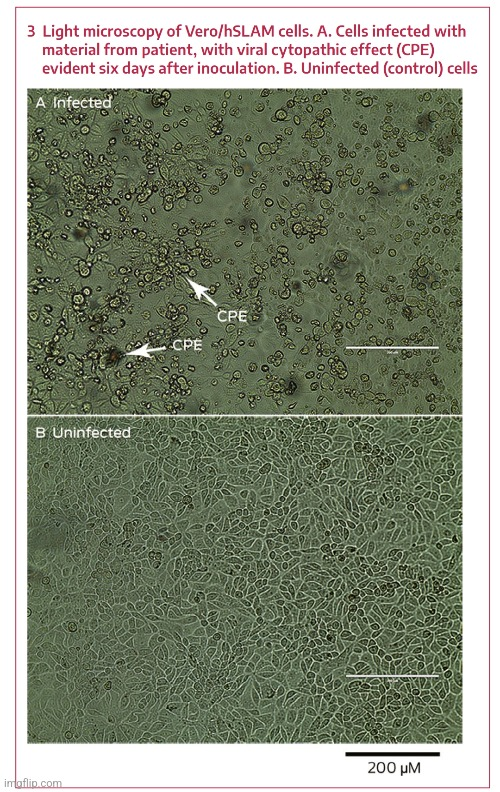
V této poslední studii od Zhoua et al. která je jedním z pilířů podvodu “SARS-COV-2”, se dozvídáme, že byl použit maketový “virus”. Co je to maketový (mock) “virus”? Zhou et al. to neuvádějí.
Vypuknutí pneumonie spojené s novým koronavirem pravděpodobného netopýřího původu.

a, b, buňky Vero E6 jsou zobrazeny 24 hodin po infekci mock virem (a) nebo 2019-nCoV (b). c, d, buňky infikované mock virem (c) nebo 2019-nCoV (d) https://www.nature.com/articles/s41586-020-2012-7.
Nebo to neřekli veřejně. V soukromé korespondenci prostřednictvím e-mailu však byly k této situaci osvětleny některé další podrobnosti. Z úžasné eseje Dr. Marka Baileyho “A Farewell to Virology” se dozvídáme, že v experimentální kultuře byla během kultivačních pokusů zdvojnásobena antibiotika, aby se dosáhlo cytopatogenního účinku v 1 z 24 kultur. Nejenže se jedná o ohromující neúspěšnost kultivace “viru”, ale přidání většího množství antibiotik do experimentální kultury zcela znehodnocuje výsledky, protože kontrolní kultura nebyla ošetřena stejně.

Vědecký podvod. Strany 41-42 brilantní eseje Dr. Marka Baileyho.
Jak je vidět na těchto příkladech, to, co bylo provedeno s kontrolními kulturami, tzv. mock-infected, není dobře definováno a je třeba to brát jako předpoklad, že obě kultury byly ošetřeny stejně po odejmutí předpokládaného “viru”. Přiznání Zhoua a spol. je však přesně důvodem, proč nemůžeme předpokládat, že s kulturami bylo zacházeno stejně, protože tomu tak zjevně nebylo. Přidání většího množství antibiotik do experimentální kultury nebylo v rámci článku nikde zmíněno. Zhou a spol. se dopustili vědeckého podvodu. U kolika dalších “virových” studií by se ukázalo, že se dopustili téhož, kdyby byli vědci tak upřímní, jako byl Xing-Lou Yang v e-mailové výměně? Podrobnosti o tom, co bylo provedeno s pokusnými infikovanými kontrolami, musí být uvedeny v každé práci, ale to se děje jen zřídka, pokud vůbec.
Pro virologii však existuje ještě větší základní problém než nedostatek špatně definovaných kontrolních vzorků mock-infection, a to i v případě, že jsou použity všechny stejné přísady. Nezapomeňte, že kontrola má eliminovat pouze jednu studovanou proměnnou, tj. předpokládané “virové” částice. Vzhledem k tomu, že se připouští, že tekutiny, které se používají pro “inokulovanou” kulturu, neobsahují pouze purifikované a izolované “virové” částice, ale celou škálu potenciálních látek, jako jsou hostitelské materiály, bakterie, houby, mikrovezikulární tělesa atd., nejsou mock-infections, kdy se do kultury nepřidávají žádné lidské tekutiny, správnou kontrolou. Správnou kontrolou by bylo použití vzorku zdravého člověka, který je zpracován přesně stejným způsobem jako tekutiny s předpokládaným “virem”. To zahrnuje přidání zdravých lidských tekutin do “virového” transportního média, které obsahuje přidané chemikálie, živiny, fetální hovězí sérum, antibiotika/antimykotika atd., protože tento krok se provádí ihned po odběru “virového” vzorku. Vynechání tekutin od zdravých osob z kontroly znehodnocuje modelovou infekci, protože v experimentální kultuře je přítomno mnoho matoucích proměnných, které v modelové infikované kultuře chybí. Ačkoli lze tedy říci, že kontrolní vzorek infikované tekutiny je kontrolou, není správnou kontrolou pro pseudovědecké experimenty s buněčnými kulturami.

Jak by měly být prováděny správné kontroly v rámci metody pseudovědecké buněčné kultury. Převzato z publikace No “Virus” Challenge. (ZDE je CZ verze)
Shrnutí:
- Studie s kontrolou (kontrolami) je navržena tak, aby bylo zajištěno, že účinky jsou způsobeny nezávislými proměnnými v experimentu
- Použití kontrol umožňuje studovat jednu proměnnou nebo faktor v daném okamžiku
- Je důležité, aby kontrolní i druhá (experimentální) skupina (skupiny) byly vystaveny stejným podmínkám kromě jedné zkoumané proměnné
- Ve vztahu k virologii by touto proměnnou byly předpokládané “virové” částice a proto je nezbytné, aby virologové používali neočištěný vzorek od nemocných lidí a proto by museli jako kontrolu použít i neočištěný vzorek od zdravého člověka
- V průběhu počátku 20. století se objevily velmi rozdílné interpretace podstaty “virů”, které se stavěly proti sobě.
- Žádná ze stran nemohla předložit experimentální důkazy pro tu či onu koncepci, kterou měli všichni badatelé akceptovat
- Zjištění, která často uváděli někteří tehdejší badatelé zabývající se “viry”, nebyla potvrzena jinými badateli jako výsledek jejich vlastních experimentů nebo pozorování nemohla být reprodukována všemi vědci, kteří s “viry” pracovali
- Často byly hlášeny opačné nálezy nebo byly zkoumané nálezy považovány za artefakty
- Každá verze interpretace fenoménu “viru” zůstávala otevřená útokům
- Fakta předkládaná odborné veřejnosti mohla být často oponenty reinterpretována ve fikce, které jako zdroje chyb uváděly závislost zjištění na podmínkách pozorování, místní situaci experimentů, výzkumnou povahu atributů vlastností atd.
- K nálezům, které sloužily k empirickému potvrzení domnělé souvislosti, se často brzy přidaly negativní nálezy, o nichž informovali jiní badatelé
- V žádném okamžiku jedna strana nenabídla přesvědčivé důvody, které by druhou stranu vedly k tomu, aby se nakonec vzdala obvinění z artefaktů
- V roce 1954 praxe tkáňových kultur, která prokazatelně přinášela stejné výsledky i ve zdravých tkáních, upadla v nemilost ve prospěch nové metody buněčných kultur Johna Franklina Enderse
- Buněčná kultura se spoléhala na vytvoření cytopatogenního efektu (CPE), tedy buněčná smrt, jako specifických pro “viry”, aby bylo možné identifikovat, že jsou v kultuře přítomny
- Přestože tato metoda byla rychle přijata jako nový zlatý standard pro “izolaci virů”, sám Enders své výsledky zpochybnil, protože pozoroval nerozlišitelnou buněčnou smrt i v kontrolních zdravých kulturách, jakou viděl u kultur s “virem” spalniček
- Podle Enderse bylo druhé agens získáno z neinokulované kultury buněk opičích ledvin
- Cytopatické změny, které vyvolal v neobarvených preparátech, nebylo možné s jistotou odlišit od “virů” izolovaných ze spalniček
- Považoval za zřejmé, že možnost setkání s takovými agens při studiích se spalničkami je třeba mít neustále na paměti
- Ke stejným závěrům dospěli i další výzkumníci v průběhu následujících pěti let, neboť stejný cytopatogenní účinek pozorovali i u svých vlastních kontrolních kultur zdravých jedinců

- Nebezpečí vnášení “virů” do tkáňových kultur s kontaminovanými médii bylo zdůrazněno nedávno
- Detekce takových “virových” infekcí může být v některých případech komplikována skutečností, že se neprojevují žádné snadno rozpoznatelné změny (tzv. úniková klauzule “virů” bez CPE).
- V roce 1953 se vědci pokusili adaptovat havajský “virus” dengue adaptovaný na myši na válečkové zkumavkové kultury opičích ledvin rhesus a došlo k výskytu neidentifikovaného agens, které vyvolalo cytopatogenní změny v kulturách opičích ledvin a nádorových epiteliálních buněk HeLa
- Tři další agens s identickými cytopatogenními vlastnostmi přešla z neinokulovaných kultur opičích ledvin připravených pro studie poliomyelitidy na kultury HeLa buněk
- Enders a Peebles oznámili získání agens z nenaočkované kultury opičích ledvin, které měly stejné cytopatogenní vlastnosti v opičích ledvinových kulturách jako jejich agens
- Byly předloženy důkazy, že zdrojem těchto látek v tkáňových kulturách opičích ledvin jsou ledviny zjevně zdravých opic a nikoli složky média
- Krátce po setkání s agens MK-D při pokusech o adaptaci “viru” dengue na kultury opičích ledvin byly v neinokulované zkumavkové kultuře 12 dní po její přípravě opět pozorovány syncytiální hmoty a vakuoláty
- Agens MK3 a MK4 bylo podobně získáno z neinokulovaných kultur připravených z ledvin různých opic rhesus
- Byly získány informace, které naznačují, že frekvence výskytu těchto agens v neočkovaných kulturách může být relativně vysoká
- Výzkumníci důsledně pozorovali prakticky ve všech kulturách připravených z různých opičích ledvin občasné malé vakuoly patrné od počátku buněčného růstu
- Výzkumníci uvedli, že tato zjištění poskytují důkaz, že tyto látky pocházejí z ledvinové tkáně opic a že zdrojem těchto látek je ledvinová tkáň, a nikoli složky média.

- Enders a Peebies pozorovali, že kultury lidské nebo opičí ledvinové tkáně inokulované vzorky od pacientů se spalničkami podléhají po jedné nebo více pasážích charakteristickým jaderným a jiným cytologickým změnám
- V pokusech s buněčnými kulturami na zarděnky, které provedli Cohen a kol., se kontroly skládaly z neočkovaných kultur a z kultur pasážovaných sériově s tekutinami z neinokulovaných zkumavek
- Přenosné agens, pravděpodobně “viry”, byly údajně izolovány jak z krve, tak z výtěru z krku jednoho pacienta a z výtoku z nosu a výtěru z krku dalších dvou pacientů, jejichž krev nebyla získána
- Ve vzorcích z dalších 5 případů nebyli zjištěni žádní původci (agens)
- Inokuly od 3 z nich byly před kultivací skladovány při teplotě 4°-6 °C po dobu 2 dnů; testy inokul od dalších 2 byly přerušeny po 3 zjevně negativních sériových pasážích a Cohen se domníval, že tyto okolnosti mohly přispět k tomu, že se u nich agens nepodařilo izolovat
- Cohen zopakoval, že Enders a Peebles a Rustigian et al. se setkali s latentními “virům podobnými” původci, kteří vyvolávají výraznou vakuolizaci a syncytiální hmoty v tkáňových kulturách opičích ledvin
- Mějte na paměti, že nepozorovali žádná “virům podobná” agens, pouze předpokládali, že musí být přítomna, protože stejný CPE byl pozorován u kontrol, které údajně žádný “virus” neobsahovaly
- Buněčná degenerace charakteristická pro tyto “opičí ledvinové agens” se často objevovala v našich kulturách a to jak v těch, které byly inokulovány vzorky od pacientů se spalničkami, tak v kontrolních; proto bylo obtížné použít cytologická kritéria pro rozpoznání spalničkových agens
- Jinými slovy, CPE byl totožný, protože nemohli rozeznat žádný rozdíl mezi CPE pozorovaným u neočkovaných kontrol a u vzorků očkovaných a nemohli použít CPE jako kritéria pro rozpoznání “viru” spalniček
- Cohen a spol. neopakovali Endersovy filtrační experimenty, ale nebyli schopni získat plísně, uzliny z králíků imunizovaných difterií nebo bakterie včetně organismů podobných pleuropneumonii a leptospiry z infikovaných tkáňových kultur, takže předpokládali, že antigeny, které vyvolaly specifické reakce ve fluorescenčních testech na protilátky a komplement-fixaci, pocházejí z “viru” spalniček
- Jinými slovy, protože nemohli použít CPE k určení případů spalniček, použili výsledky nespecifických protilátek, o kterých předpokládali, že jsou specifické pro “virus” spalniček

- Jak popsali Enders & Peebles a později Rustigian et al. a Cohen el al. cytopatické změny podobné těm, které způsobuje “virus” spalniček, lze pozorovat i v neinokulovaných kulturách tkáně opičích ledvin
- Tyto změny jsou pravděpodobně způsobeny virům podobnými agens, tzv. pěnovými agens, které se zřejmě často vyskytují v ledvinových buňkách zdánlivě zdravých opic
- Tento druhý krok degenerace vedl k nahromadění buněčných zbytků s kruhovými zrnitě vypadajícími útvary, které měly buď zcela hladký, nebo zvrásněný okraj (buněčné zbytky s kruhovým útvarem určitě zní hodně jako “virus” 🤔)
- Opičí ledvinové “viry” nebo “pěnové agens”, mohou dát vzniknout buněčným degeneracím, které jsou mikroskopicky nerozlišitelné od těch, které způsobuje “virus” spalniček
- Rozhodli, že z tohoto důvodu mají cytologické projevy při studiu spalniček omezenou hodnotu a že jsou zapotřebí další kritéria pro stanovení identity kultivovaných původců
- Ve studii prezentované v tomto článku byl “virus” spalniček izolován v pěti z devíti výplachů krku odebraných do 24 hodin po propuknutí exantému
- Předpokládá se, že důvodem neúspěšné izolace u jednoho pacienta bylo to, že bezprostředně před kloktáním zvracel
- Ve zbývajících třech případech nelze nabídnout žádné zjevné vysvětlení negativních výsledků, ale domnívali se, že by to mohlo být způsobeno tím, že všechny tyto izolace byly provedeny pomocí výtěrů z krku u malých dětí, které byly velmi rozrušené, a odběr vzorků se tak obtížně prováděl
- “Virus” byl získán z krve pouze v jednom z osmi pokusů provedených do 24 hodin po vzniku vyrážky, což bylo málo ve srovnání s výsledky, které získali Enders & Peebles, kteří získali “virus” z krve čtyř z pěti pacientů
- Uvažovalo se o možnosti, že neúspěch při získání “viru” u 8 ze 13 vyšetřovaných pacientů by mohl být způsoben necitlivostí konkrétních použitých buněk vůči “viru” spalniček, ale tato teorie byla opuštěna
- Konstatovali, že pozorování uvedená v této práci dobře souhlasí s předpokladem, že izolované agens je příčinou spalniček
- Všechny pokusy izolovat “virus” později než 24 hodin po vzniku vyrážky selhaly
- Byly popsány cytopatické projevy pozorované u tkáňových kultur infikovaných “virem” spalniček i u neinokulovaných zkumavek
- Intranazální a orální podání materiálu z pozdních pasáží jednoho z izolovaných původců dvěma opicím rodu Rhesus vedlo u jednoho ze zvířat k výrazné vyrážce podobné spalničkám a u obou opic se vyvinuly protilátky proti naočkovanému kmeni

- Hull a kol. uvedli, že širší využití techniky kultivace buněk pro izolaci, udržování a studium “virů” vedlo k objevu mnoha dosud neznámých cytopatogenních agens
- Různí výzkumníci objevili řadu “virových agens”, která jsou přítomna v tkáních a výkalech člověka a nižších živočichů a která se vzpírají detekci jinými metodami než tkáňovými kulturami
- Během výroby a testování vakcíny proti poliomyelitidě byly v našich laboratořích a v dalších laboratořích, které se podílely na tomto programu, pozorovány statisíce opičích ledvinových kultur připravených z tisíců opic a objevily se i četné filtrovatelné, přenosné cytopatogenní agens jiné než “poliovirus”
- Hull uvedl, že ačkoli se tato agens pravděpodobně dostala do kultur s buňkami opičích ledvin, zůstává vzdálená možnost, že některá představují kontaminanty z lidských zdrojů, koňského séra, živného média nebo jiných roztoků používaných při přípravě kultur
- Izolovaní agens byli označováni jako “opičí viry” (S.V.), a to až do doby, než se podařilo určit jednoznačné spojení s nějakým jiným hostitelem nebo identifikaci
- Jedním z nich byl S.V.13, což je “krajkovitý nebo pěnivý” agens, který se často vyskytuje v kulturách opičích ledvinových buněk, o nichž se hovoří ve výše uvedených studiích.
- Pro S.V. 1 bylo použito 1 700 zkumavek připravených ze směsi ledvin 4 zjevně zdravých opic rodu cynomolgus.
- Po 6 až 8 dnech inkubace se kultury jevily jako uspokojivé a byly použity k titraci “polioviru” a po dalších 6 dnech inkubace byl v některých kulturách pozorován typ destrukce buněk za koncovým bodem “polioviru”, který byl netypický pro poškození způsobené “poliovirem”
- Jinými slovy, delší inkubační doba vedla k dalším cytopatogenním účinkům, které (subjektivně) nebyly považovány za způsobené obrnou, takže bylo rozhodnuto, že je musel způsobit jiný “původce”
- Toto atypické poškození bylo pozorováno u 17 % kultur
- Opice byly inokulovány intracerebrálně, intramuskulárně a v některých případech i jinými cestami všemi “opičími viry” a s výjimkou S.V.12 a S.V.15 nebyly pozorovány žádné známky klinického onemocnění ani hrubých či histopatologických změn
- Opice inokulované intracerebrálně vysoko titrovými neředěnými “viry” S.V.12 a S.V.15 podlehly během 4 až 6 dnů po inokulaci.
- Stejné dva “viry” však při intramuskulární inokulaci opicím nevyvolaly klinické onemocnění ani hrubé či histopatologické změny
- Je vidět, že předpokládané S.V. nevykazovaly žádné známky patogenity, s výjimkou S.V. 12 a 15, které po injekci do mozku opice usmrtily, ale po injekci do svalů nic nezpůsobily, takže se nejedná o “viry”
- Nebyla zjištěna žádná rozpoznatelná choroba u opic, kterým byly tyto “viry” aplikovány.
- Žádný z těchto opičích “virů” neinfikoval embryonální vajíčka, což bylo potvrzeno smrtí embrya.
- Skupiny dospělých a sajících myší byly inokulovány intracerebrálně a intraperitoneálně každým “virem” a přesto nebyly pozorovány žádné známky onemocnění ani úhynu myší
- Potkani a králíci, kteří byli inokulováni živým “virem” za účelem výroby antiséra, rovněž nepodlehli infekci žádným z těchto agens
- Byly popsány “izolace” a vlastnosti 8 zřejmě nových “virů”, avšak nebyl předložen žádný jednoznačný důkaz a předpokládalo se, že tito původci jsou pravděpodobně opičího původu
- Nebylo zjištěno, jaká onemocnění, pokud vůbec nějaká, by mohly u opic vyvolávat
- Jinými slovy, výzkumníci nemohli vyvolat onemocnění těmito “původci” nalezenými v očkovaných i neočkovaných kulturách na základě identifikace pomocí CPE a tudíž se nejednalo o “viry”
- Mnoho z těchto agens bylo získáno z normálních nebo neočkovaných kontrolních kultur
- Hull et al. opakují, že bylo popsáno osm zřejmě nových submikroskopických, filtrovatelných, cytopatogenních agens získaných v tkáňových kulturách tkáně ledvin opic rhesus a cynomolgus
- Mezi žádným z těchto původců a přirozeně se vyskytujícími chorobami opic nebo jiných zvířat nebyla zjištěna žádná jednoznačná souvislost

- Dr. Stefan Lanka provedl vlastní kontrolní pokusy s buněčnými kulturami, při nichž byl podvod CPE odhalen (CZ verze textu ZDE)
- V kontrolním pokusu č. 1 byly normální buňky kultivovány s normálním živným médiem a pouze malým množstvím antibiotik a ani první, ani pátý den nebyl zjištěn žádný CPE; buňky pokračovaly v normálním, zdravém růstu
- V kontrole č. 2 byly normální buňky opět pěstovány na normálním živném médiu a malém množství antibiotik, ale tentokrát bylo k obohacení média přidáno 10% fetálního telecího séra a přesto buňky v kultuře rostly normálně a to jak první, tak pátý den
- Kontrola č. 3 zahrnovala změnu živného média na “minimální živné médium” – což znamená snížení procenta fetálního telecího séra z obvyklých 10 % na 1 %, což snižuje množství živin dostupných pro růst buněk a tím je stresuje – a ztrojnásobení koncentrace antibiotik
- Pátý den pokusu se objevil charakteristický CPE, který “prokázal” existenci a patogenitu “viru” – až na to, že v žádném okamžiku nebyl do kultury přidán patogenní “virus”
- Tento výsledek může znamenat pouze to, že CPE byl důsledkem způsobu provedení kultivačního pokusu a nikoliv “viru”
- Kontrola č. 4 je stejná jako kontrola č. 3 s tím rozdílem, že k této kultuře byl přidán roztok čisté RNA z kvasinek, což vedlo ke stejnému výsledku jako ve sloupci 3, což opět dokazuje, že příčinou CPE je technika kultivace – a nikoli “virus”
- Podle technického bulletinu o buněčných kulturách, když se kultury samovolně zničí bez zjevné příčiny a nelze nalézt žádné důkazy o běžných biologických kontaminantech, jsou často obviňovány kryptické “viry”
- Bylo uvedeno, že je to nešťastné, protože skutečnou příčinou destrukce kultury může být něco jiného, možná mykoplazma nebo chemický kontaminant a v důsledku toho zůstane neodhalena, aby se stala vážnějším problémem
- Mezi další faktory, které jsou údajně schopny vyvolat CPE, patří např:
- Bakterie
- Améba
- Paraziti
- Exosomy
- Antibiotika
- Antimykotika
- Chemické kontaminanty
- Stáří buňky
- Stres způsobený prostředím
- Experimenty na buněčných kulturách mají zahrnovat podrobné pokusné kontroly (mock-infections), ale málokdy se tak děje.
- S mock-infekcemi se má zacházet stejně jako s experimentálními kulturami, jen bez “viru”
- Někdy se v mock-infikovaném vzorku používá “nevirulentní kmen“.
- V klíčové studii “SARS-COV-2”, kterou provedl Zhou a kol., byla antibiotika v experimentální kultuře během kultivačních pokusů zdvojnásobena, aby se dosáhlo cytopatogenního účinku pouze v 1 z 24 kultur a tato informace byla v publikované studii vynechána
- Bez ohledu na to, mock-infekce nejsou správnou kontrolou, protože by měla být vyloučena pouze studovaná proměnná (“virus”)
- Vzhledem k tomu, že použité nečištěné tekutiny obsahují také další hostitelské a cizorodé materiály, kontaminanty a znečišťující látky, je vyloučeno i mnoho matoucích proměnných, pokud se s maketovou infikovanou kulturou nepoužije lidský vzorek
- Vhodnou kontrolou pro tyto pseudovědecké pokusy s buněčnými kulturami by tedy bylo použití vzorku ze zdravého hostitele, se kterým se zachází úplně stejně jako se vzorkem z nezdravého hostitele, s “virovými” transportními médii a vším ostatním

Pokud chce být virologie považována za skutečnou vědu, musí se řídit vědeckou metodou. To znamená, že experimenty a kontroly, které jsou obhajovány jako “zlatý standard” důkazů pro tento obor, musí být navrženy v souladu s touto metodou. Virologie má však své základy pevně zakořeněné v pseudovědě, protože nikdy neexistoval žádný přirozeně pozorovaný jev, na němž by byla virologie postavena. “Virus” je imaginární konstrukt vysněný v hlavách výzkumníků, kteří pravidelně nenacházeli bakteriální příčinu onemocnění a kteří pak předpokládali, že v tekutinách nemocných lidí musí být obsaženo něco jiného, menšího a nepozorovatelného. Stejně jako nejlepší lovci Yettiho hledali virologové tyto mytologické entity po celá desetiletí a nepodařilo se jim nalézt přímé důkazy o jejich přítomnosti ani přímo v tekutinách nemocného hostitele, ani prostřednictvím různých groteskních tkáňových kultur a nelidských pokusů na zvířatech. Pokusy prokázat existenci fiktivních stvoření měly sestupnou tendenci a v padesátých letech 20. století bylo vše ztraceno a obor byl připraven zemřít přirozenou smrtí.
Situace se však pro virologii obrátila k lepšímu, když John Franklin Enders v roce 1954 představil svou metodu podvodných buněčných kultur a nakonec umírající obor oživil. Virologové nyní dokázali zabíjet buňky kombinací různých toxinů v Petriho misce a vytvářet tak malebná umělecká díla, která se objevují ve všech médiích jako zobrazení těchto mytologických bytostí. Následný důkaz a usmrcení buňky, označovaný jako cytopatogenní účinek, byl nově definován jako účinek specifický pro “virus”, aby se pro neexistující nezávislou proměnnou vytvořila neplatná závislá proměnná. Místo přímých důkazů existence těchto entit byla vytvořena nová forma nepřímých důkazů, aby bylo možné oblbnout masy. Virologie úspěšně vykouzlila vlastní laboratorně vytvořené proměnné, s nimiž si hrála, místo aby prokázala příčinu a následek studiem jakýchkoli reálných jevů.
I když se obor držel metody buněčných kultur, aby získal pseudovědecké důkazy potřebné k udržení virologie při životě, okamžitě se objevil závažný problém, na který upozornil Enders, vynálezce této techniky a který popsal jako svědectví svých prvních experimentů. CPE, z něhož byl obviňován “virus”, byl pozorován jak během “infikovaných” kultur, tak i v jeho “neinfikovaných” kontrolních kulturách. To měl být smrtelný úder pro virologii, protože bylo jasné, že toxické experimentální uspořádání způsobilo účinek, který byl přičítán neviditelnému “viru”, jak ukázaly kontroly. Jako by to nestačilo, různí další výzkumníci po zbytek padesátých let hlásili stejná zjištění, protože totožný CPE, z něhož byl obviňován “virus”, byl pozorován u kontrolních kultur zdravých opic, které údajně žádné “viry” neobsahovaly. Někteří výzkumníci připustili, že tento efekt nelze použít k tvrzení o přítomnosti “viru” a místo toho se pak spoléhali na stejně podvodné výsledky protilátek, aby podpořili svá tvrzení o “viru”. Jiní se pokoušeli tvrdit, že zdravé kontrolní kultury obsahovaly “virům podobné” látky, ale při pokusech prokázat patogenitu pomocí těchto tekutin neuspěli a tak místo toho prokázali, že nic, co by splňovalo definici “viru”, nebylo přítomno. Navíc se ukázalo, že stejný cytopatogenní účinek způsobuje i mnoho jiných faktorů než předpokládané “viry”, např. bakterie, améby, použitá antibiotika, stáří buňky atd. Virologie úspěšně vyvrátila sama sebe.
Protože podrobné popisy kontrolních kultur prokázaly, že “virus” za pozorovaný CPE nemůže, mnoho následujících prací buď přestalo podrobně popisovat, co se s kontrolami dělo nebo se prostě kontrolám vyhnulo úplně. Kontroly známé jako “mock-infections”, pokud se dnes ve studiích vyskytují, jsou špatně definované a v některých případech se ukázalo, že jsou zcela podvodné, jako například v práci Zhou et al., která s kontrolní kulturou (mock-infection) zacházela jinak než s experimentální kulturou, čímž znehodnotila výsledky zásadní práce “SARS-COV-2”. I když jsou tyto mock-infekce určitou formou kontroly, lze je jen stěží považovat za řádnou kontrolu, protože z rovnice nevylučují pouze částice “viru”. Vyloučeny jsou také ostatní potenciální organismy, kontaminující látky, chemické látky atd. v tekutinách, které byly přidány do “virového” vzorku a které jsou následně vynechány z mock-infekčních kultur, které neobsahují žádné tekutiny od lidí. Nelze tedy říci, že by se v mock-infected kulturách měnila pouze jedna zkoumaná proměnná, protože z rovnice je odstraněno mnoho proměnných. Proto mock-infection bez srovnatelného lidského vzorku bez předpokládaného “viru” není správnou kontrolou.
Pokud chce být virologie skutečnou vědou, musí se vrátit k rýsovacímu prknu. Virologové se musí pokusit pozorovat přirozený fenomén, u kterého lze nezávislou proměnnou (tj. příčinu; “virus”) pozorovat v přírodě. To přinejmenším znamená, že musí najít částice, o nichž se domnívají, že jsou “viry”, přímo v tekutinách nemocného hostitele a určit způsob, jak tyto částice oddělit od všeho ostatního v tekutinách. Virologové musí určit platnou závislou proměnnou (tj. studovaný účinek; příznaky nemoci), aby mohli stanovit testovatelnou a falzifikovatelnou hypotézu. Teprve poté bude možné navrhnout platný experiment, včetně stanovení vhodných kontrol. Tento postup nebyl nikde proveden, aby Enders vytvořil metodu buněčných kultur. A protože buněčná kultura není platným vědeckým experimentem, který by dodržoval tento postup, jsou všechny kontroly vědecky neplatné. Je tedy opravdu zbytečné se dohadovat, zda virologie používá kontroly, protože její metody jsou postaveny na pseudovědeckém základě. Můžeme se přít o to, aby virologové zavedli to, co by vzhledem k pseudovědeckým parametrům jejich experimentu bylo správnými kontrolami, ale je jasné, že i přes nedostatek používání správných kontrol, virologie již dávno vyvrátila sama sebe.
Je načase zaměřit se místo toho na jádro problému, na nedodržování vědecké metody. Nejlepší kontrolou ze všech je, když nás virologové seznámí s tím, jak jejich obor dodržuje vědeckou metodu. Jaké přírodní jevy byly pozorovány? Jak virologové identifikovali a stanovili platnou IV (nezávislou proměnnou)? Co je to DV (závislá proměnná)? Jaká hypotéza vedla k vytvoření metody buněčných kultur a jak byla stanovena? Jak byla tato metoda buněčných kultur ověřena, aniž by byl nejprve úspěšně vyčištěn a izolován “virus”, aby se zjistilo, jak jej pěstovat? Jak mohou virologové ospravedlnit použití laboratorně vytvořeného účinku k tvrzení o příčině, kterou lze pozorovat až po provedení experimentu, když je to protiklad vědecké metody? Chtít po virologii, aby ukázala, jak dodržuje vědeckou metodu, je kontrola, při které pokaždé selže. A to je jediná kontrola, která je potřeba.

Zdroj Přeložil: Vladimír Bartoš V průběhu let se vedlo mnoho diskusí o tom, zda virologie dodržuje vědeckou metodu, či nikoliv, přičemž většina těchto diskusí se zaměřovala na nedostatek řádných kontrol. Spor se soustředil na to, zda kontroly, které se někdy používají, ale jsou zřídka popsány, známé jako falešné…